
Les yeux composés et les ommatidies
La vision des odonates est complexe, comme pour tous les insectes aux yeux composés.
Quand on clique sur la photo ci-dessus, on voit correctement les ommatidies ; ce sont les unités élémentaires, qui composent l’œil de la libellule. On en compte jusqu’à 30.000 chez les Aeshnidae (Anax junius a exactement 28,672 ommatidies dans chaque œil à maturité ; Land and Nilsson 2002) ce qui constitue le record pour le monde des insectes(1). Leurs yeux, les plus gros de tous les insectes, leur permettent de voir pratiquement dans toutes les directions… sauf directement en arrière.
Ces ommatidies ressemblent à toutes celles présentes chez les insectes et forment une structure conique à section hexagonale, la partie la plus large du cône est à la surface de l’œil recouverte d’une lentille (cornée, cristallin…), la partie la plus étroite, profonde, est une structure photosensible appelée rhabdome dont s’échappe un axone qui relayé par un ganglion atteint le cerveau.
L’image résultante peut se comparer à un pixel et l’image globale est recomposée au niveau du cerveau environ 200 fois par seconde ; en conséquence, si vous invitez une libellule au cinéma, choisissez un film tourné en 200 images secondes, sinon notre standard humain (24 ou 30 I/s) lui paraîtra d’un autre âge, complètement saccadé… 🙂
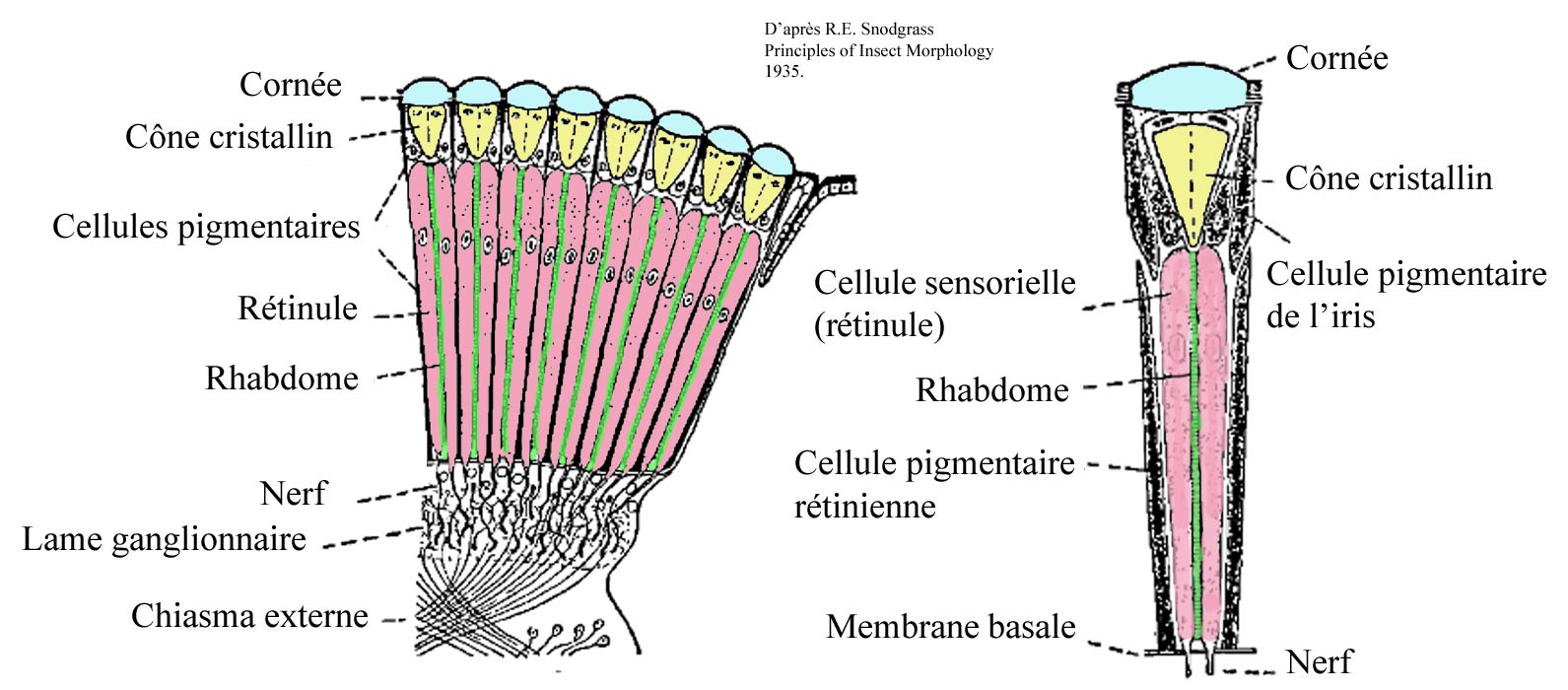
Les yeux des odonates ne sont pas mobiles comme les nôtres, mais leur conformation leur permet de voir presque « partout à la fois » sans bouger la tête, et sans doute ultérieurement de « se concentrer » sur l’image qui nécessite le traitement le plus urgent.

Les pseudo-pupilles
Cette formation d’ommatidies accolées est responsable d’un phénomène de pseudo-pupilles visible chez tous les insectes, mais plus facilement observable pour les odonates en raison de la grande taille de leurs yeux et de la couleur parfois claire de ceux-ci. Leurs yeux semblent montrer des pupilles sombres, parfois multiples qui donnent l’impression de suivre le déplacement de l’observateur.

Quand le regard de l’observateur tombe dans l’axe d’une ommatidie (ou de plusieurs très voisines) il perçoit un point noir dû au fait, que dans cet axe, la lumière n’est pas réfléchie par les cellules pigmentées qui la tapisse mais « absorbée » par la zone photo sensible profonde. Mais il ne s’agit en aucun cas d’une véritable pupille au sens du diaphragme de l’œil des mammifères.
Vision des couleurs
On a longtemps douté que les odonates aient une vision en couleur, mais ils laissent très loin derrière notre vision trichromatique basée sur l’expression de 3 gènes codant 3 opsines (protéines réagissant à l’énergie lumineuse) ; les odonates en possèdent de 15 à 33, 20 par exemple pour le Sympetrum frequens, dont 16 opsines destinées à la perception des couleurs. Ces opsines sont spécifiquement sensibles, chacune, a une certaine fourchette de longueurs d’ondes, certaines sensibles aux ondes courtes (SW), d’autres aux ondes longues (LW).
Anax parthenope fait partie des champions puisqu’il compte 33 gènes pour coder les opsines dont 8 pour les courtes longueurs d’ondes et 21 pour les longues, et bien sûr tous les odonates « voient » les UV et le plan de polarisation de la lumière, ce dernier élément joue certainement une part importante dans le contrôle du vol (2).
Les odonates voient donc parfaitement les couleurs, bien mieux que nous, et ils possèdent des récepteurs spécialisés pour les analyser très finement.
Spécialisation régionale des yeux
Comme on le voit très bien sur la photo ci-dessous (en cliquant …) les ommatidies ne sont pas de la même taille sur toute la surface de l’œil. Elles sont plus larges sur la partie supérieure de l’œil, à peine discernables sur la partie inférieure. Cette distinction morphologique correspond évidemment à une différenciation fonctionnelle.

Les larges ommatidies permettent le passage d’un grand nombre de photons et la réduction de la diffraction à travers une lentille plus large a pour conséquence une image plus nette.
Cette répartition régionale des ommatidies s’accompagne d’une répartition spécifique, quantitative et qualitative des opsines entre la partie haute et basse de l’œil. Ainsi, pour les anisoptères, on note une forte expression des gènes codant les opsines pour les ondes courtes (SW) et très peu pour les ondes longues (LW) dans la partie supérieure de l’œil. L’inverse est vrai pour la partie inférieure.
On peut, comme l’ont fait les auteurs du document (2) trouver une justification écologique et comportementale à cette répartition ; la lumière solaire venant d’en haut est de forte intensité et biaisée vers les longues ondes, la lumière de l’environnement, venant d’en bas, réfléchie, tend à être de plus faible intensité et déportée vers les longues ondes.
Cela correspond parfaitement aux conditions rencontrées par un anisoptère perché sur le bord d’un étang qui surveille son petit territoire en explorant et surveillant vers l’avant et le bas, d’où provient la lumière réfléchie, et scrutant (en même temps !) le ciel pour y déceler d’éventuelles proies (ou prédateurs).
Enfin, il a été montré (3) que le bord supérieur de l’œil comportait une région spécialisée dans le traitement de la lumière polarisée et cette zone intervient certainement dans le contrôle de la navigation de l’insecte.
-1 – All the better to see you with: a review of odonate color vision with transcriptomic insight into the odonate eye, Org Divers Evol (2012) 12:241–250.
-2- Extraordinary diversity of visual opsin genes in dragonflies, Futahashi et al., 2015.
-3- Meyer, E.P. & Labhart, T. Cell Tissue Res (1993) 272: 17. https//doi.org/10.1007/BF00323566

